
El entrenamiento intenso activa la síntesis de proteínas, pero solo si existe la nutrición adecuada para apoyarla. Si lees el artículo que John Meadows y yo escribimos llamado Maximizar la síntesis de proteínas, o si has leído algo de lo que T Nation ha estado diciendo sobre el tema durante años, probablemente estés familiarizado con el concepto de "la ventana anabólica" y la importancia de nutrición peri-entrenamiento.
Lo que sucede a nivel celular en las horas posteriores al entrenamiento es predictivo de ganancias a largo plazo. Aproveche esta "ventana anabólica" y crecerá como nunca antes. Lo echo de menos constantemente y, bueno, buena suerte.
Obtener los macronutrientes adecuados en el momento adecuado es la clave, pero las macros son solo una parte del panorama general. Un aspecto importante, pero a menudo pasado por alto, de la síntesis de proteínas musculares es el volumen celular. El volumen celular no es solo cosmético; Es el principal impulsor del transporte de aminoácidos, y también trabaja entre bastidores para activar la síntesis de proteínas y suprimir la degradación de proteínas.
Un músculo lleno / voluminoso es un músculo anabólico. Si bien sabemos desde hace más de 20 años que la hinchazón celular inhibe la degradación de proteínas y estimula la síntesis de proteínas en ciertas células (1-3), hasta hace poco, el mecanismo que vincula el volumen celular con la síntesis de proteínas era un misterio.
Lo que sabemos ahora es que la síntesis de proteínas está controlada por la enzima mTOR, que es activada por estrés mecánico, factores de crecimiento y leucina.
Si bien los tres son importantes para el estímulo de entrenamiento, la señalización de mTOR también depende del volumen celular.(4) Esto es especialmente importante en el músculo esquelético, donde la voluminización celular activa la síntesis de glucógeno y proteínas e inhibe la degradación de proteínas.(5, 6)
El avance científico que condujo a la conexión entre el volumen celular y la síntesis de proteínas se produjo en 2005, cuando un grupo de científicos descubrió que se necesita más que solo leucina para activar mTOR; también se requiere glutamina.(7)
Esto fue una sorpresa. Aunque la glutamina se considera un aminoácido "condicionalmente esencial" que limita la degradación de proteínas durante un trauma o estrés severo, nunca se había relacionado con la activación de mTOR.
La glutamina era necesaria para la absorción de leucina y la voluminización celular, las cuales son necesarias para activar la síntesis de proteínas. Los autores continuaron demostrando que la glutamina celular agotamiento no solo da como resultado una reducción del volumen celular, sino que también reduce la capacidad de la leucina para activar la síntesis de proteínas.(7)
Este descubrimiento fue enorme, porque proporcionó un vínculo directo entre la glutamina, la voluminización celular y la síntesis de proteínas. Por primera vez, se demostró que la glutamina es necesaria para la activación de la síntesis de proteínas por la leucina.
Si bien este estudio sugirió que la glutamina es una pieza muy importante del rompecabezas que une el volumen celular y la síntesis de proteínas, el mecanismo exacto no se resolvió hasta 2009, cuando Nicklin et al. descubrió que la exportación de glutamina está acoplada a la importación de leucina y la activación de mTOR.(8)
Para que la leucina entre en la célula, hay un período inicial de "carga de glutamina."Esto también atrae agua, lo que aumenta el volumen celular. Después de la fase de "carga de glutamina", la glutamina se exporta fuera de la célula a cambio de la importación de leucina.
Nicklin y col. También descubrió que los niveles de glutamina celular limitan la velocidad de activación de la síntesis de proteínas por la leucina. Cuando las células se trataron simultáneamente con glutamina y una mezcla de EAA que contenía leucina, la activación de la síntesis de proteínas se retrasó durante 60 minutos. Cuando estas mismas células se "precargaron" con glutamina, la síntesis de proteínas se activó dentro de 1-2 minutos después de que las células recibieron leucina.
Este resultado fue importante porque explicó el tiempo de retraso para la activación de la síntesis de proteínas por leucina en este modelo experimental.
Estos resultados finalmente arrojan algo de luz sobre la maquinaria celular que regula el transporte de aminoácidos y cómo se acopla al control de la síntesis de proteínas.
Este trabajo debe interpretarse con cierto grado de cautela, sin embargo. Una advertencia importante para estos estudios fue que se realizaron in vitro (i.mi. en cultivos celulares) donde la regulación o síntesis de proteínas es mucho más simple. Las células musculares son capaces de producir glutamina según sea necesario a partir de otros aminoácidos, y el "agotamiento de la glutamina" en este modelo de cultivo celular no es representativo de situaciones más fisiológicas in vivo.
Las altas tasas de síntesis de proteínas no se pueden mantener indefinidamente en el tejido muscular con o sin suplementos de glutamina. Sin embargo, la glutamina se puede utilizar para respaldar estratégicamente la síntesis de proteínas al optimizar la voluminización celular durante el período posterior al entrenamiento.

Las células están muy ocupadas y hay muchos canales iónicos unidos a la membrana y proteínas transportadoras que regulan el tráfico dentro y fuera de la célula. Hay dos clases de transportadores de aminoácidos en particular que son importantes aquí: los transportadores de aminoácidos del "Sistema L" y del "Sistema A" están más estrechamente relacionados con la señalización de mTOR y la síntesis de proteínas.(8-10)
La actividad de los transportadores del Sistema A y del Sistema L está acoplada, lo que permite que la leucina y los otros BCAA se absorban en la célula.(11) Los transportadores del Sistema L son responsables de la entrada de leucina y otros BCAA a cambio de la salida de glutamina.
Sin embargo, los transportadores del sistema A funcionan a través de un mecanismo diferente, donde la glutamina se acopla a la absorción de sodio.(12, 13) El acoplamiento entre la absorción de sodio y los transportadores de aminoácidos del Sistema L / Sistema A se denomina Transporte Activo Terciario (TAT para abreviar). Es TAT el que finalmente impulsa la leucina dentro de la célula, lo que conduce a la activación de mTOR y la síntesis de proteínas.(11)
Puede ver cómo funciona TAT en la siguiente figura:
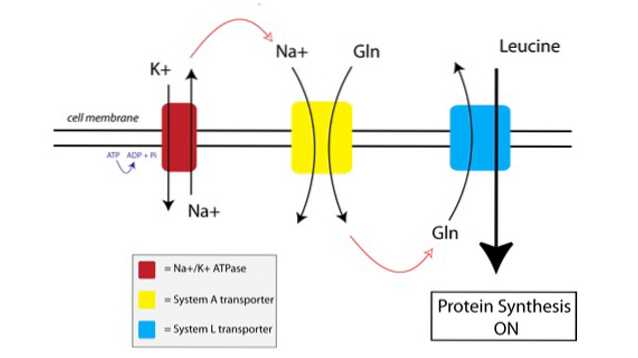
Primero, una bomba unida a la membrana llamada bomba de ATPasa de sodio-potasio (Na + / K + ATPasa, rojo en la figura anterior) usa energía del ATP para mover el sodio fuera de la célula, contra su gradiente de concentración.
El aumento de la concentración de sodio fuera de la célula se acopla a la importación de glutamina por el transportador del Sistema A (amarillo en la figura). El influjo de glutamina y sodio en la célula también atrae más agua, lo que hace que la célula se hinche. Esto pone a la célula en un estado anabólico, preparando la maquinaria de síntesis de proteínas para su activación.
Cuando la glutamina alcanza niveles suficientemente altos dentro de la célula, se activan los transportadores del Sistema L (azul en la figura), que transportan la glutamina fuera de la célula a cambio de la absorción de leucina. La entrada de leucina en la célula es el desencadenante de la síntesis de proteínas.
Si bien esta ha sido una gran lección de bioquímica hasta ahora, el descubrimiento de TAT no solo es importante para los biólogos celulares. Ahora que sabemos cómo se acopla el volumen celular al transporte de aminoácidos y la síntesis de proteínas, podemos diseñar varias estrategias nutricionales para maximizar el proceso cuando cuenta, durante el período crítico posterior al entrenamiento.
La síntesis de proteínas depende totalmente de la hidratación celular: si está un poco deshidratado, la capacidad de recuperarse de un entrenamiento intenso se ve totalmente comprometida. Obtener mucha agua es una obviedad aquí, pero el agua por sí sola no es suficiente.
Los electrolitos como el sodio, el potasio, el cloruro y el fosfato también funcionan como "osmolitos" porque atraen agua hacia la célula. Después de una intensa sesión de entrenamiento, necesitamos agua, aminoácidos y electrolitos para maximizar el proceso de voluminización celular que impulsa la síntesis de proteínas.
El sodio, magnesio, calcio, potasio, fosfato y cloruro (por nombrar algunos) son todos importantes aquí. En un nivel mínimo (a menos que lo esté haciendo bajo el consejo de su médico), no evite el sodio antes o después del entrenamiento. Si tiene poco sodio, su bombeo del entrenamiento será casi inexistente y el sodio es necesario para la absorción de glutamina.
Para eliminar las conjeturas, Surge® Workout Fuel y Plazma ™ están diseñados con proporciones ideales de electrolitos para respaldar el volumen celular y la síntesis de proteínas.
La captación de glutamina en la célula provoca voluminización celular, lo que prepara a las células musculares para la síntesis de proteínas. Como se mencionó, un músculo lleno / voluminoso es un músculo anabólico. Además de impulsar el transporte de aminoácidos, la voluminización celular también aumenta la síntesis de glucógeno e inhibe la degradación de proteínas.(4-6)
La síntesis de proteínas es suprimida por el agotamiento de la glutamina, lo que tiene enormes implicaciones para los atletas que entrenan duro. Después de una intensa sesión de entrenamiento, se genera una respuesta inflamatoria, que permite que las células inmunitarias entren en el tejido muscular golpeado para comenzar el proceso de reparación / reconstrucción.(14)
La glutamina es absorbida tan rápidamente por las células inmunitarias que se considera el "combustible del sistema inmunológico".”(15) No es sorprendente que se haya demostrado que el entrenamiento intenso provoca el agotamiento de la glutamina plasmática.(16-18)
Por esta razón, los requerimientos de glutamina aumentan en el período posterior al entrenamiento, donde la respuesta inmune local puede competir por la disponibilidad de glutamina para preparar las células musculares para la absorción de aminoácidos y la síntesis de proteínas.
Precarga las células con glutamina también pueden reducir potencialmente el "tiempo de retraso" asociado con la activación de la síntesis de proteínas por leucina. Si aún no lo está haciendo, tome 10-15 g de glutamina o péptidos de glutamina inmediatamente después del entrenamiento. Debido a que los BCAA son otro sustrato favorito para la síntesis de glutamina muscular y también se ha demostrado que aumentan la producción de glutamina muscular (19-21), los BCAA y la leucina también son útiles durante el período previo al entrenamiento para ayudar a maximizar la producción de glutamina endógena.
Nota del editor: Todo el protocolo Plamza / MAG-10, con su nutrición previa al entrenamiento, el entrenamiento y el posentrenamiento, cumple con las recomendaciones del autor para cargar los músculos con BCAA y leucina.
Recientemente, se descubrió que el consumo de EAA aumenta la expresión de los transportadores de aminoácidos del Sistema A y del Sistema L.(9) Es importante destacar que esto sucede en el "nivel postranscripcional", es decir, en el nivel de síntesis de proteínas, donde los ARNm existentes se traducen en proteínas.
Compare esto con "denovo"Expresión de proteínas - donde puede llevar 16 horas o más sintetizar, procesar y transportar nuevos ARNm - la activación postranscripcional de la síntesis de proteínas puede ocurrir en minutos u horas, lo que permite que las células aumenten rápidamente el nivel de proteínas particulares cuando sea necesario.
De repente, tenemos más incentivos para tener un plan de nutrición sólido durante el período de entrenamiento: la ingesta de EAA durante los períodos de pre y peri entrenamiento da buenos resultados después del entrenamiento al aumentar la expresión de los transportadores de aminoácidos, preparando la célula para una absorción máxima de aminoácidos. y activación de la síntesis de proteínas.
Los aislados o hidrolizados de proteínas de rápida absorción como Mag-10® Protein Pulsing Protocol ™ o Plazma ™ durante los períodos de pre y peri-entrenamiento son ideales aquí.
La insulina es la hormona más anabólica del cuerpo. Junto con la activación directa de la síntesis de proteínas, la insulina también aumenta la translocación de los transportadores de aminoácidos del Sistema A a la membrana celular.(22)
Esto significa que la insulina hace que se muestren más transportadores del Sistema A en la membrana celular, listos para conducir más glutamina al interior de la célula. Más glutamina conduce a más volumen celular, lo que lleva más leucina a la célula, lo que finalmente conduce a una mayor síntesis de proteínas.
Mientras que los EAA aumentan la expresión de los transportadores AA, es la señal de insulina la que les permite mostrarse en la superficie celular, listos para transportar nuevos aminoácidos a la célula.
Esta es otra razón más por la que los carbohidratos antes y durante el entrenamiento son una buena idea, a menos que esté en modo de pérdida de grasa extrema: la insulina aumenta la capacidad de transporte celular de aminoácidos.
Los carbohidratos aumentan los niveles de insulina, pero ciertos aminoácidos también se pueden usar para potenciar la liberación de insulina. La glutamina es un poderoso activador de las hormonas "incretinas", que hacen que las células productoras de insulina del páncreas sean más sensibles a la glucosa.(23) La glicina también potencia la liberación de insulina a través de un mecanismo diferente.
Aunque los carbohidratos después del entrenamiento por sí solos aumentarán los niveles de insulina, la combinación de estos aminoácidos que potencian la insulina con carbohidratos sobrecargará su páncreas para una liberación aún mayor de insulina. Si bien es bueno mantener los niveles de insulina en el lado más bajo la mayor parte del tiempo, el aumento de los niveles de insulina en el período del peri-entrenamiento maximiza el transporte de aminoácidos, el volumen celular y la síntesis de proteínas al mismo tiempo que suprime la degradación de proteínas.
El tipo de entrenamiento anaeróbico intenso que se necesita para desarrollar una gran cantidad de músculos de calidad conduce a una producción considerable de lactato, lo que disminuye el pH muscular. Esto conduce a una fatiga y debilidad muscular tempranas, pero también ciertos transportadores de aminoácidos, incluido el Sistema A, son inhibidos por un pH bajo.(13)
Cuando el pH muscular es bajo, la captación de aminoácidos se reduce, lo que suprime la activación de mTOR de la síntesis de proteínas.(24) También se ha demostrado que la inhibición del pH bajo de los transportadores de aminoácidos del Sistema A aumenta la degradación de proteínas.(25)
Aquí es donde entra en juego la beta-alanina. El aumento de los niveles de carnosina muscular actúa como un amortiguador ácido natural, extendiendo el umbral anaeróbico al limitar la disminución del pH muscular debido al entrenamiento.
La beta-alanina tiene otra función importante: ayudar a mantener la síntesis de proteínas y ponerla en línea más rápido después de un entrenamiento intenso al evitar la atenuación del transporte de aminoácidos.
Para aumentar los niveles de carnosina muscular, tome dos tabletas de Beta-7 ™ tres veces al día.
Durante las sesiones de entrenamiento intenso, se reduce la síntesis de proteínas y se activa la degradación de proteínas. Esto es inevitable para cualquier levantador de entrenamiento duro. Sin embargo, la medida en que podamos minimizar los efectos catabólicos del entrenamiento y lo más rápido que podamos volver al "modo anabólico" durante el período posterior al entrenamiento determina en última instancia la eficacia con la que nos recuperaremos y creceremos.
La sincronización de los macronutrientes es importante, pero es un medio para lograr un fin. El volumen celular es el principal impulsor del transporte de aminoácidos y la síntesis de proteínas. Al comprender cómo ocurre el transporte de aminoácidos y cómo está regulado por el volumen celular, podemos obtener más leucina en las células musculares destruidas más rápido, avivando así el fuego anabólico y, en última instancia, conduciendo a mejores ganancias.
Las estrategias anteriores son efectivas, prácticas y se basan en las últimas investigaciones científicas. Úselos como plantilla para llevar su progreso de entrenamiento al siguiente nivel.
Espero sus preguntas sobre LiveSpill!



Nadie ha comentado sobre este artículo todavía.